Sexual Reproduction in Flowering Plants
Pre-Fertilisation Events
- Several hormonal and structural changes result in the development of a flower.
- Inflorescences bear the flower buds, and then the flowers.
- Flowers are the reproductive parts of a plant.
- In the flowers, the androecium (male reproductive part) and the gynoecium (female reproductive part) develop.
Androecium
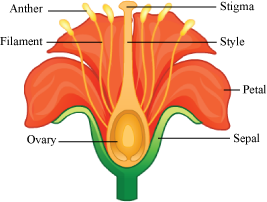
- The androecium consists of whorls of stamen.
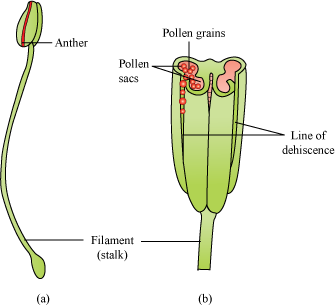
- The stamen consists of the filament (long and slender stalk) and anther (bilobed structure).
- Filament is attached to the thalamus or to the petal.
- Anther:
- A typical anther is bilobed and each lobe is dithecous (consists of two theca).
- Theca are separated by a longitudinal groove running lengthwise.
- The microsporangia are located at the corners, two in each theca. They further develop to form pollen sacs, which contain the pollen grains.
- Structure of microsporangium
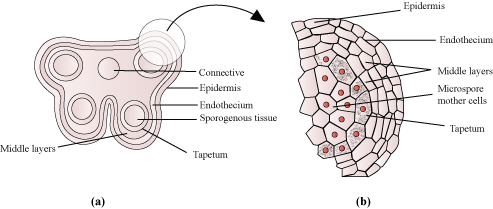
- The microsporangium is surrounded by four wall layers (epidermis, endothecium, middle layers, and tapetum).
- The outer three layers are protective and help in dehiscence of anther to release the pollen grains. The tapetum provides nourishment to the developing pollen grains.
- In the young anther, the sporogenous tissue forms the centre of each microsporangium.
Microsporogenesis
- It is the process of formation of microspore from PMC (Pollen Mother Cells).
- As development occurs in the anther, the sporogenous tissue undergoes meiosis to form microspore tetrad.
- Each cell of sporogenous tissue has capacity to give rise to a tetrad. Hence, each cell is a potential pollen or PMC.
- As the anther matures, the microspores get detached from each other and develop into pollen grains.
Pollen grains
- Represent the male gamete and are spherical, having a two-layered wall:
- Exine (outer) − Hard layer made of sporopollenin, which is extremely resistant and can withstand high temperatures, acidic and alkaline conditions, and enzymes
- Intine (inner) − Thin and continuous layer made up of cellulose and pectin
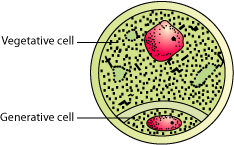
- Mature pollen grain contains two cells:
- Vegetative cell − Large with irregular nucleus, contains food reserves
- Generative cell − Small and floats in the cytoplasm of the vegetative cell
- In 60% of the angiosperms, pollen grains are shed at 2-celled stage while in others generative cell undergoes mitosis to form two male gametes (3-celled stage).
- The viability of pollen grains after they are shed depends upon temperature and humidity. It ranges from 30 minutes to few months.
Gynoecium and Formation of Female Gametophyte
- The gynoecium represents the female reproductive part of a flower.
- It may be mono-carpellary (one pistil) or multi-carpellary (many pistils). In multi-carpellary, the pistils may be fused in one (syncarpous) or free (apocarpous).
- Each pistil consists of:
- Stigma − Receives the pollen grains
- Style − Elongated, slender part below the stigma
- Ovary − Bulged basal part containing the placenta, which is located inside the ovarian locule (cavity)
- The placenta contains the megasporangia or ovules.

Megasporangium
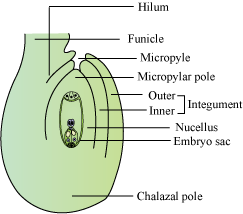
- The ovule is attached to the placenta by the funicle. The junction of the ovule and the funicle is called hilum.
- Each ovule has one or two protective layers, called integuments, which cover the rest of the ovule, except for a small opening called micropyle.
- The chalaza lying on the opposite side of the micropyle end represents the basal part of the ovule.
- Nucellus is present within the integuments and contains reserved food. The embryo sac or female gametophyte is located within the nucellus.
Megasporogenesis
- The megaspore mother cell (MMC) gets converted into megaspores by the process of megasporogenesis.
- The MMC is large and contains a dense cytoplasm and a prominent nucleus. It undergoes meiosis to produce four megaspores.
Female Gametophyte
- In most flowering plants, only one megaspore is functional while the other three degenerate.
- The single functional megaspore develops into the female gametophyte. This kind of development is called monosporic development.
- The nucleus of the functional megaspore divides mitotically to form 2 nuclei, which move towards the opposite ends, forming a 2-nucleate embryo sac. Two more mitotic divisions ensue, leading to the formation of 4-nucleate and 8-nucleate embryo sacs.
- After the 8-nucleate stage, the cell walls are laid down and the typical female gametophyte (embryo sac) gets organised.
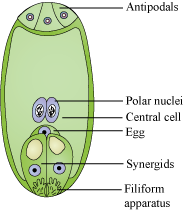
- Six of the 8-nuclei get surrounded by the cell wall and the remaining two, called polar nuclei, are situated below the egg apparatus in the large central cell.
- Three of the six cells are placed at the micropylar end and constitute the egg apparatus (2 synergids + 1 egg cell).
- The synergids have special thickenings at the micropylar end. These are together called the filiform apparatus. It helps in leading the pollen tubes into the synergids.
- Three cells are at the chalazal end, and are called antipodal cells.
- A typical angiosperm female gametophyte is 7-celled and 8-nucleated at maturity.
Pollination
- It is the process of transfer of pollen grains from the anther to the stigma.
- Depending on the source of pollen, pollination can be divided as follows:
- Autogamy − It is the transfer of pollen grains from the anther to the stigma of the same flower. Autogamy requires the anther and the stigma to lie close. It also requires synchrony in the pollen release and stigma receptivity.
Plants like Viola, Oxalis, etc., produce two kinds of flowers—chasmogamous flowers (with exposed anther and stigma) and cleistogamous flowers (which do not open at all and only autogamy occurs). - Geitonogamy − It is the transfer of pollens from the anther of one flower to the stigma of another flower in the same plant. Genetically, it is similar to autogamy, but it requires pollinating agents.
- Xenogamy − It is the transfer of pollen grains from the anther to the stigma of a different plant. Pollination causes genetically different types of pollens to be brought to a plant.
Agents of Pollination
- Plants use air, water (abiotic agents) and animals (biotic agents) for pollination.
- Pollination by wind
- It is the most common form of abiotic pollination.
- Plants possess well-exposed stamens and large, feathery stigma.
- Pollens should be light and non-sticky to be carried easily by winds.
- Wind-pollinated flowers often have single ovule in the ovary and numerous flowers packed in an inflorescence.
- It is common in grass.
- Pollination by water
- It is rare in flowering plants, except for some aquatic plants like Vallisneria and Hydrilla.
- In most water-pollinated plants, the pollen grains are long and ribbon-like, and are protected from wetting by mucilaginous covering.
- In a majority of water plants like water hyacinth and water lily, flowers emerge above the water level and are pollinated by insects.
- Pollination by animals
- Majority of flowering plants use butterflies, bees, wasps etc., for pollination.
- Most of the insect-pollinated flowers are large, colourful, fragrant, and contain nectar to attract the animal pollinators. These are called floral rewards.
- Floral reward can be in the form of providing safe places to lay eggs (example: the tallest flower, Amorphophallus)
- A symbiotic relationship exists between the plant, Yucca and its pollinator moth. The moth is dependent on the plant since the moth deposits its eggs in the locule of the ovary of the plant, and in return, the plant is pollinated by the moth.
- The pollen grains are sticky and get stuck to the body of the pollinator.
Out Breeding Devices
- Repeated self pollination leads to inbreeding depression.
- Plants have developed methods to prevent self pollination. Autogamy is prevented by following ways:
- Pollen release and stigma receptivity not coordinated
- Different positioning of the anther and the stigma
- Production of unisexual flowers
- Ways to prevent both autogamy and geitonogamy:
- Presence of male and female flowers on different plants, such that each plant is either male or female (dioecy).
- This mechanism is present in several species of papaya.
Pollen−Pistil Interactions
- Pollination does not always ensure the transfer of compatible pollens.
- Hence, the pistil has the ability to recognise the right type of pollen to promote post- pollination events.
- If the pollen is of the wrong type, the pistil prevents pollen germination.
- This interaction is mediated by chemical components of the pollen and the pistil.
- Pollen−pistil interaction is a dynamic process involving pollen recognition, followed by promotion or inhibition of the pollen.
- The pollen tube reaches the ovary and enters the ovule through the micropyle. Then, through the filiform apparatus, it reaches synergids. In this way, the pollen tube grows.
Artificial Hybridisation & Double Fertilisation
Artificial Hybridisation
- It is a method to improve crop yield.
- In this method, it is essential to ensure that the right kinds of pollen grains are used, and the stigma is protected from unwanted pollen grains. It is achieved by:
- Emasculation − The anther is removed from the bud if the female parent bears bisexual flowers.
- Bagging − The emasculated flower is covered by a bag so as not to allow contamination of the stigma by unwanted pollen grains.
- When the stigma of the bagged flower becomes receptive, the collected pollen grains are dusted onto the stigma, and then the flower is rebagged.
- If the female parent is unisexual, emasculation is not necessary. In this case, the female bud is directly bagged, and when the stigma turns receptive, suitable pollen grains are dusted onto it so as to allow germination.
Double Fertilisation
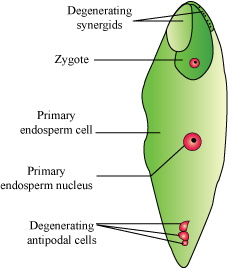
- When the pollen grains fall on the stigma, the pollen tube enters one of the synergids and releases two male gametes.
- One of the male gametes moves towards the egg cell and fuses with it to complete the syngamy to form the zygote.
- The other male gamete fuses with the two polar nuclei and forms triploid primary endosperm nucleus (PEN). This is termed as triple fusion.
- Since two kinds of fusion—syngamy and triple fusion—take place, the process is known as double fertilisation, and is characteristic of flowering plants.
- After triple fusion, the central cell becomes the primary endosperm cell (PEC).
- The primary endosperm nucleus gives rise to the endosperm, while the zygote develops into the embryo.
Post-Fertilisation Events
It includes development of endosperm and embryo, and maturation of ovules into seeds and ovaries into fruits.
Formation of Endosperm
- The endosperm develops before the embryo because the cells of the endosperm provide nutrition to the developing embryo.
- The primary endosperm nucleus repeatedly divides to give rise to free nuclei. This stage of development is called free nuclear endosperm.
- Cell wall formation occurs next, resulting in a cellular endosperm.
- The endosperm may be either fully consumed by the growing embryo (as in pea and beans) or retained in the mature seed (as in coconut and castor).
Development of Embryo
- The embryo develops at the micropylar end of the embryo sac where the zygote is situated.
- The zygote gives rise first to the pro-embryo, and then to the globular, heart-shaped, mature embryo.
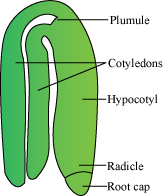
- A typical dicot embryo consists of an embryonal axis and two cotyledons.
- The portion of the embryonal axis above the level of cotyledons is called epicotyl. It contains the plumule (shoot tip). The portion below the axis is called hypocotyl. It contains the radicle (root tip). The root tip is covered by the root cap.
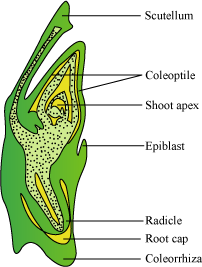
- In a monocot embryo, there is only one cotyledon. In grass, it is known as the scutellum, and is situated at one side of the embryonal axis. At its lower end, the embryonal axis has the radicle and the root cap enclosed in the coleorrhiza.
- The epicotyl lies above the level of the scutellum, and has the shoot apex and leaf primordia enclosed in hollow structures called coleoptiles.
Seeds and Fruits
Development of Seeds
- It is the last stage of sexual reproduction in angiosperms.
- Seeds are the fertilised ovules that are developed inside a fruit.
- A seed consists of:
- Seed coat
- Cotyledons
- Embryonal axis
- Seeds may be albuminous (endosperm present; as in wheat and maize) or non-albuminous (endosperm absent; since it is consumed by the growing embryo; as in pea and beans).
- Some seeds such as black pepper and wheat have remnants of nucellus known as perisperm.
- The integuments of ovules harden to form the seed coat, and the micropyle facilitates the entry of oxygen and water into the seed.
- As it loses moisture, the seed may enter dormancy, or if favourable conditions exist, it germinates.
Development of Fruits
- The ovary of a flower develops into a fruit.
- The walls of the ovary transform into the walls of the fruit (pericarp).
- Fruits may be fleshy, as in mango and orange, or can be dry, as in groundnut and mustard.
- In some plants, floral parts other than the ovary take part in fruit formation, as in apple and strawberry. In these, the thalamus contributes to fruit formation. Such fruits are called false fruits. Fruits that develop from the ovary are called true fruits.
- Some fruits develop without fertilisation, and are known as parthenocarpic fruits (example: banana).
Apomixis and Polyembryony
- Some plants produce seeds without fertilisation. This process of seed formation is known as apomixis.
- Apomixis is a form of asexual reproduction mimicking sexual reproduction.
- In some species, apomixis occurs as the diploid egg cell is formed without meiosis, and develops into embryo without fertilisation.
- In some varieties of citrus and mango, the nucellus cells divide and protrude into the embryo sac to develop into embryos. In such cases, each ovule may contain several embryos and this condition is called polyembryony.
- Apomixis is important for producing hybrid varieties of fruits and vegetables, and also for increasing crop yield multifold.